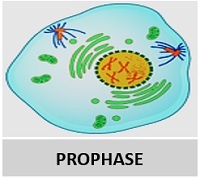
Pada prometafase (profase lanjut) kromosom memadat di dalam amplop inti dan aster serat muncul di bagian luar kromosom.
Ketika selaput nukleus telah hilang, sebuah spindel terbentuk di prometafase. Serat gelendong terdiri dari bundel mikrotubulus yang memancar dari ujung yang berlawanan dan disebut sebagai kutub sel.
Kromosom kemudian bermigrasi ke bidang ekuatorial tempat mereka menempel pada salah satu serat spindel.
Dalam sel hewan, pembentukan gelendong terjadi oleh sentrosom, yang terdiri dari sentriol kembar pada sudut kanan yang dikelilingi oleh bahan amorf.
Sentrosom adalah pusat pengorganisasian mikrotubulus utama selama interfase. Replikasi sentrosome dalam fase G1 dan S akhir dan pasangan dapat diamati tepat di luar amplop nuklir.
Namun, tanaman yang lebih tinggi tidak memiliki sentrosom yang khas meskipun nukleasi dan dinamika mikrotubulus mereka menunjukkan bahwa tanaman memiliki aktivitas pusat pengatur mikrotubulus (MTOC) yang bergantung pada siklus sel.
Inisiasi polimerisasi mikrotubulus dalam sel biasanya terjadi pada situs nukleasi spesifik yang disebut sebagai MTOC. Pada sebagian besar tanaman tingkat tinggi, inisiasi mitosis ditandai oleh dua peristiwa berturut-turut yang melibatkan populasi mikrotubulus yang berbeda.
Ini adalah produksi pita preprofase dan perkembangan gelendong bipolar. Mikrotubulus sitoplasma dari tanaman yang lebih tinggi memancar dari permukaan nukleus menuju korteks sel.
Ini adalah salah satu aspek khusus dari sitoskeleton tanaman dibandingkan dengan jenis sel lainnya. Permukaan nuklir pabrik dapat terdiri dari aktivitas MTOC dan ini dapat mewakili salah satu faktor penting dalam pengendalian inisiasi mitosis.
Ada bukti eksperimental untuk mendukung peran permukaan nukelus tumbuhan sebagai situs nukleasi mikrotubulus. Ketika mitosis dimulai dan kromosom menjalani kondensasi, penggabungan tubulin telah terbukti meningkat pada permukaan nuklir sel endosperma Haemanthus dalam profase, berpotensi sebagai kontrol nukleasi yang bergantung pada siklus sel.
Peristiwa kromosom penting dari prometafase adalah perlekatan kromosom pada gelendong dan gerakannya menuju pusat gelendong.
Lampiran kromosom pada spindel terjadi di kinetokor, yang mengandung protein untuk perlekatan kromatid. Perincian amplop nuklir memungkinkan kinetokor untuk menempel pada mikrotubulus spindel.
Prometafase merupakan tahap proses pembelahan sel, perantara antara profase dan metafase. Prometafase ditandai dengan interaksi kromosom dalam divisi dengan mikrotubulus yang akan bertugas memisahkannya.
Baik dalam mitosis dan meiosis, prometafase terjadi, tetapi dengan karakteristik yang berbeda.
Tujuan yang jelas dari semua pembelahan sel adalah untuk menghasilkan lebih banyak sel. Untuk mencapai ini, sel awalnya harus menduplikasi konten DNA-nya; yaitu, meniru itu. Selain itu, sel harus memisahkan kromosom ini sehingga tujuan khusus dari setiap divisi sitoplasma terpenuhi.
Pada mitosis, jumlah kromosom yang sama dari sel induk dalam sel anak. Pada meiosis I, pemisahan antara kromosom homolog. Pada meiosis II, pemisahan antara kromatid saudara. Artinya, pada akhir proses, dapatkan empat produk meiotik yang diharapkan.
Sel menangani mekanisme rumit ini melalui penggunaan komponen khusus seperti mikrotubulus. Ini diselenggarakan oleh sentrosom di sebagian besar eukariota. Di tempat lain, sebaliknya, seperti tanaman unggul, jenis lain dari pusat organisasi mikrotubulus bekerja.
Mikrotubulus
Mikrotubulus adalah polimer linear dari protein tubulin. Mereka campur tangan di hampir semua proses seluler yang melibatkan perpindahan beberapa struktur internal. Mereka adalah bagian integral dari sitoskeleton, silia dan flagela.
Dalam kasus sel tumbuhan, mikrotubulus juga berperan dalam organisasi struktural internal. Dalam sel-sel ini, mikrotubulus membentuk semacam permadani yang melekat pada wajah bagian dalam membran plasma.
Struktur mikrotubulus, yang mengendalikan pembelahan sel tumbuhan, dikenal sebagai organisasi kortikal mikrotubulus. Pada saat pembelahan mitosis, misalnya, mereka runtuh menjadi cincin pusat yang akan menjadi situs masa depan dari pelat pusat, di bidang di mana sel akan membelah.
Mikrotubulus terdiri dari alpha-tubulin dan beta-tubulin. Dua subunit ini membentuk heterodimer, yang merupakan unit struktural dasar filamen tubulin.
Polimerisasi dimer mengarah ke pembentukan 13 protofilamen dalam organisasi lateral yang menimbulkan silinder berongga.
Silinder berongga dari struktur ini adalah mikrotubulus, yang dengan komposisinya sendiri menunjukkan polaritas. Artinya, satu ujung dapat tumbuh dengan menambahkan heterodimer, sedangkan ujung lainnya dapat dikurangi. Dalam kasus terakhir, alih-alih memanjang ke arah itu, mikrotubulus menyusut.
Mikrotubulus nukleat (mis. Mulai mempolimerisasi) dan mengorganisasikannya menjadi pusat pengorganisasian mikrotubulus. Ini dikaitkan dengan sentrosom selama pembelahan dalam sel hewan.
Pada tumbuhan yang lebih tinggi, yang tidak memiliki sentrosom, pusat pengorganisasian mikrotubulus hadir di situs analog, tetapi dibentuk oleh komponen lain. Pada silia dan flagela, ini terletak pada dasarnya pada struktur motorik.
Perpindahan kromosom selama pembelahan sel dilakukan melalui mikrotubulus. Ini memediasi interaksi fisik antara sentromer kromosom dan pusat pengorganisasian mikrotubulus.
Dengan reaksi depolimerisasi yang ditargetkan, kromosom metafase akhirnya akan bergerak menuju kutub sel pembagi.
Prometafase mitosis
Segregasi kromosom mitosis yang benar adalah pemisahan yang memastikan bahwa setiap sel anak menerima pelengkap kromosom yang identik dengan sel induk.
Ini berarti bahwa sel harus memisahkan setiap pasangan kromosom yang digandakan menjadi dua kromosom yang terpisah. Artinya, ia harus memisahkan kromatid sister dari setiap pasangan homolog dari semua komplemen kromosom sel induk.
Mitosis terbuka
Dalam mitosis terbuka, proses hilangnya selaput nukleus adalah ciri khas prometafase. Ini memungkinkan satu-satunya penghalang antara MOC dan sentromer kromosom menghilang.
Dari MOC, untaian mikrotubulus yang panjang dipolimerisasi, yang memanjang ke arah kromosom. Setelah menemukan centromere, polimerisasi berhenti dan kromosom yang melekat pada pusat pengorganisasian mikrotubulus diperoleh.
Pada mitosis kromosomnya berlipat ganda. Oleh karena itu, ada juga dua sentromer, tetapi masih bersatu dalam struktur yang sama. Ini berarti bahwa pada akhir proses polimerisasi mikrotubulus kita akan memiliki dua di antaranya per kromosom duplikat.
Satu filamen akan mengikat sentromere ke pusat pengorganisasian mikrotubulus, dan yang lainnya ke kromatid yang menempel pada pusat pengorganisasian mikrotubulus berlawanan dengan yang pertama.
Mitosis tertutup
Dalam mitosis tertutup, prosesnya hampir identik dengan yang sebelumnya, tetapi dengan perbedaan besar; amplop nukleus tidak hilang. Oleh karena itu, pusat pengorganisasian mikrotubulus bersifat internal dan dikaitkan dengan amplop inti internal melalui lamina nuklir.
Dalam mitosis semi-tertutup (atau semi-terbuka), selubung inti hanya menghilang pada dua titik yang berlawanan di mana mitosis pusat pengorganisasian mikrotubulus ada di luar nukleus.
Ini berarti bahwa dalam mitosis ini mikrotubulus menembus ke dalam nukleus untuk memobilisasi kromosom dalam langkah-langkah setelah prometafase.
Metafase meiosis
Karena meiosis melibatkan produksi empat sel ‘n’ dari satu sel ‘2n’, harus ada dua divisi sitoplasma. Mari kita lihat seperti ini: Pada akhir metafase I akan ada empat kali lebih banyak kromatid daripada sentromer yang terlihat di bawah mikroskop.
Setelah pembelahan pertama, akan ada dua sel dengan kromatid dua kali lebih banyak daripada sentromer. Hanya pada akhir pembelahan sitoplasma kedua semua sentromer dan kromatid akan individual. Akan ada sentromer sebanyak kromosom.
Protein utama untuk interaksi interkromatik yang kompleks ini terjadi dalam mitosis dan meiosis adalah cohesin. Tetapi ada lebih banyak komplikasi pada meiosis daripada pada mitosis. Oleh karena itu, tidak mengherankan bahwa kohesi meiosis berbeda dari kohesi mitosis.
Kohesi memungkinkan kohesi kromosom selama proses kondensasi mitosis dan meiotik. Selain itu, mereka memungkinkan dan mengatur interaksi antara kromatid sister di kedua proses.
Tetapi dalam meiosis mereka juga mempromosikan sesuatu yang tidak terjadi dalam mitosis: kawin antar homolog, dan sinapsis-sinapsis berikutnya. Protein ini berbeda dalam setiap kasus. Kita dapat mengatakan bahwa meiosis tanpa cohesin yang khas tidak akan mungkin terjadi.
Meiosis I
Secara mekanis, interaksi sentromer / pusat pengorganisasian mikrotubulus adalah sama di semua pembelahan sel. Namun, pada prometafase I meiosis I, sel tidak akan memisahkan kromatid saudara seperti pada mitosis.
Sebaliknya, tetrad meiosis memiliki empat kromatid dalam satu set sentromer ganda. Dalam struktur ini ada hal lain yang tidak ada dalam mitosis: kiasma.
Kiasma, yang merupakan penyatuan fisik antara kromosom homolog, adalah apa yang membedakan sentromer mana yang harus dipisahkan: kromosom homolog.
Ini adalah bagaimana dalam Prometafase I koneksi terbentuk antara sentromer homolog dan COM pada kutub yang berlawanan dari sel.
Meiosis II
Prometafase II ini lebih mirip dengan prometafase mitosis daripada prometafase meiotik I. Dalam hal ini, pusat pengorganisasian mikrotubulus akan “meluncurkan” mikrotubulus pada duplikat sentromer kromatid saudara.
Dengan demikian, dua sel dengan kromosom individu akan diproduksi sebagai produk dari satu kromatid dari masing-masing pasangan. Oleh karena itu, sel dengan komplemen kromosom haploid spesies akan berasal.